Pankreasa sok se sastoji od dva sekretorni proizvoda od ključne važnosti za pravilno varenje: probavnih enzima i bikarbonata. Enzima sintetiziraju i izlučuju iz acinusnih ćelija egzokrine, dok bikarbonata se luči iz epitelnih ćelija obloge mali gušterače kanala.
Probavnih Enzima
Pankreas luči veličanstveni baterija enzima koji kolektivno imaju sposobnost da se smanji gotovo sve probavljive makromolekule u oblike koji su u stanju, ili skoro u stanju da apsorbuje. Tri glavne grupe enzimi su kritični za efikasno probavu:
1. Proteaze
Probavu proteina pokreće pepsin u želucu, ali najveći dio proteina varenje zbog pankreasa proteaze. Nekoliko proteaza se sintetiziraju u pankreasu i luči u lumen tankog crijeva. Dva glavna pankreasa proteaza su tripsin i himotripsin, koji se sintetiziraju i pakirani u sekretorni vezikule kao neaktivni proenzimi tripsinogena i chymotrypsinogen.
Kao što ste mogli predvidjeti, proteaza su prilično opasni enzimi imati u ćelijama, a pakovanje od neaktivnog prekursora je način za ćelije sigurno rukovanje ovih enzima. Sekretornu vezikule također sadrži tripsin inhibitor koji služi kao dodatnu zaštitu treba da su neki od tripsinogena se aktivirati na tripsina; sljedeći egzocitoza ovaj inhibitora razredili i postaje neefikasan – pin je iz granata.
Kada se tripsinogena i chymotrypsinogen pušten u lumen tankog crijeva, moraju se pretvoriti u njihove aktivne oblike, kako bi se varenje proteina. Tripsinogena se aktivira enzim enterokinase, koji je ugrađen u crevne sluzokože.
Kada se formira tripsina aktivira chymotrypsinogen, kao i dodatne molekule tripsinogena. Neto rezultat je prilično eksplozivan izgled aktivnih proteaze jednom pankreasa sekreta do tankog crijeva.
Tripsina i himotripsin varenje proteina u peptida i peptida u manje peptide, ali oni ne mogu probaviti proteine i peptide jedne amino kiseline. Neki od drugih proteaza iz pankreasa, na primjer carboxypeptidase, imaju tu mogućnost, ali konačna probavu peptida u amino kiseline je u velikoj mjeri učinak peptidaze na površini malih crijevnih epitelnih ćelija. Više o tome kasnije.
2. Pankreasa Lipaza
A glavna komponenta masti je triglicerida, ili neutralan lipida. A triglicerida molekula ne može se direktno apsorbira preko sluznice crijevnih. Umjesto toga, prvo se mora vari u 2-Monoglyceride i dvije slobodne masne kiseline. Enzim koji obavlja ovu hidroliza je pankreasa lipaze, koji se dostavlja u lumen creva kao sastavni pankreasa soka.
Dovoljne količine žučnih soli također mora biti prisutan u lumen crijeva kako bi lipaze se efikasno varenje ishrani triglicerida i za rezultat masne kiseline i monogliceridi da se upije. To znači da normalno probavu i apsorpciju masti je kritično ovisi o sekreta iz oba pankreasa i jetre.
Pankreasa lipaza je nedavno bio u centru pažnje kao meta za upravljanje pretilosti. The orlistat droga (Xenical) je pankreasa lipaze inhibitor koji ometa probavu triglicerida i na taj način smanjuje apsorpciju masti. Klinička ispitivanja podržavaju tvrdnju da inhibira lipaze može dovesti do značajnog smanjenja tjelesne težine kod nekih pacijenata.
3. Amilaza
Glavni ishrane ugljenih hidrata za mnoge vrste je škrob, skladištenje oblik glukoze u biljkama. Amilaza (tehnički alfa-amilaza) je enzim koji hidrolizuje skrob na maltozu (glukoze-glukoze disaharid), kao i trisaccharide maltotriose i mali fragmenti branchpoints zove granica dekstrina. Glavni izvor amilaze u svim vrstama je pankreasa sekreta, iako amilaza je također prisutan u pljuvački nekih životinja, uključujući i ljude.
Ostali pankreasa Enzimi
Pored proteaza, lipaza i amilaza, pankreas proizvodi niz drugih probavnih enzima, uključujući ribonukleaze, deoxyribonuclease,’elatinaza i elastaza.
Bikarbonata i vode
Epitelnih stanica u pankreasa kanali su izvor bikarbonata i vode koji luči gušterača. Bikarbonata je osnova i kritične važnosti za neutralizaciju kiseline dolaze u tankom crijevu iz želuca. Mehanizam u osnovi bikarbonata sekrecija je u suštini isti kao i za sekreciju kiseline po parijetalni ćelija u želucu i zavisi od enzima karboanhidraze. U gušterače kanala ćelijama, bikarbonata se luči u lumen kanala i stoga u gušterače sok.
Stajati na Streetcorner i pitati ljude da li znaju šta je insulin, a mnogi će odgovarati: “Da li to nije ima neke veze sa šećerom u krvi?” Zaista, to je tačno, ali takav odgovor je pomalo kao da kažete “Mozart? Zar nije bio neki muzičar?”
Insulin je ključni igrač u kontroli posrednika metabolizam, a veliku sliku je da organizuje korištenje goriva za bilo skladištenje ili oksidacije. Kroz ove aktivnosti, inzulin ima duboke efekte na oba ugljenih hidrata i metabolizam lipida, i značajan uticaj na proteina i metabolizam minerala. Shodno tome, poremećaj u insulina signalizaciju imaju široko rasprostranjena i razarajuće efekte na mnoge organe i tkiva.
Inzulin receptor i mehanizam djelovanja
Kao i receptore za druge proteina hormona, receptor za inzulin je ugrađen u plazma membrani. Inzulin receptor se sastoji od dva alfa podjedinice i dvije beta podjedinice povezane disulfid obveznica. Alfa lanci su u potpunosti ekstracelularne i kuća vezivanja insulina domena, dok je povezan beta lance prodiru kroz plazma membranu.
Inzulin receptor tirozin kinaze. Drugim riječima, ona funkcioniše kao enzim koji prenosi fosfatne grupe iz ATP u tirozin ostatke na intracelularni ciljne proteine. Vezivanje inzulina u alfa podjedinica uzrokuje beta podjedinica fosforilišu sebe (autofosforilacije), čime se aktivira katalitičku aktivnost receptora. Aktivirani receptor zatim fosforilizuje niz intracelularni proteina, što mijenja njihove aktivnosti, čime se stvaraju biološkog odgovora.
Nekoliko intracelularni proteini su prepoznati kao fosforilacije supstrata za insulin receptor, najbolje studirao od kojih je inzulin receptor supstrat 1 ili IRS-1. Kada je IRS-1 aktivira fosforilacije, mnogo stvari se dešavaju. Između ostalog, IRS-1 služi kao neka vrsta docking centar za regrutaciju i aktiviranja drugih enzima koji su na kraju posreduje efekte insulina. Detaljniji pogled na ove procese predstavljen je u poglavlju o insulina signala.
Insulin i metabolizam ugljikohidrata
Glukoza je oslobođen od dijetetskih ugljenih hidrata kao što su škrob ili saharoze hidrolizom u tankom crijevu, a zatim se apsorbira u krv. Povišene koncentracije glukoze u krvi stimuliše oslobađanje inzulina, a inzulin djeluje na ćelije thoughout telu da stimuliše unos, korištenje i skladištenje glukoze. Efekte insulina na metabolizam glukoze variraju u zavisnosti od ciljne tkiva. Dva važna efekti su:
1. Inzulin omogućava ulazak glukoze u mišićne, masno i nekoliko drugih tkiva. Jedini mehanizam kojim ćelije mogu uzeti glukozu je po olakšana difuzija kroz porodice heksoza transportera. U mnogim tkiva – mišića kao najbolji primjer – glavni transporter za preuzimanje glukoze (tzv GLUT4) koji je dostupan na membranu kroz djelovanje inzulina.
Kada se koncentracija inzulina niska, transporteri GLUT4 glukoze su prisutni u citoplazmatski vezikule, gdje su beskorisni za transport glukoze. Vezivanje inzulina na receptore na takve ćelije brzo dovodi do fuzije onih vezikule sa plazma membrane i ubacivanje transportera glukoze, dajući ćelije sposobnost da efikasno preuzme glukoze. Kada se više ne zauzeta krvi pada inzulina i inzulin receptora, transportera glukoze se reciklira natrag u citoplazmi.
Ovdje treba napomenuti da postoje neke tkiva koja ne zahtijevaju inzulin za efikasno uzimanja glukoze: važni primjeri su mozga i jetre. To je zato što ove ćelije ne koriste GLUT4 za uvoz glukoze, već, još jedan transporter koji nije insulin zavisni.
2. Inzulin stimulira jetru za skladištenje glukoze u obliku glikogena. Veliki dio glukoze apsorbira iz tankog crijeva se odmah zauzimaju hepatocita, što ga pretvoriti u skladište polimer glikogena.
Inzulin ima nekoliko efekata u jetri koji stimulišu sintezu glikogena. Prvo, ona aktivira enzim heksozakinazom koji fosforilizuje glukoze, to zarobivši unutar ćelije. Bezmalo, inzulin djeluje na inhibira aktivnost glukoza-6-fosfataza. Inzulin također aktivira nekoliko enzima koji su direktno uključeni u sintezu glikogena, uključujući phosphofructokinase i glikogen sintaze. Neto efekt je jasna: kad je u izobilju opskrbu glukoze, inzulina “govori” jetru da banke kao mnogo toga je moguće koristiti kasnije.
3. A dobro poznati učinak inzulina je smanjenje koncentracije glukoze u krvi, što bi trebalo da ima smisla s obzirom na mehanizme gore opisano. Još jedna važna uzeti u obzir da, kao što je koncentracija glukoze u krvi padne, lučenje inzulina prestaje. U absense insulina, bulk ćelija u organizmu postaju u stanju da preuzmu glukozu, i početi prelazak na korištenje alternativnih goriva poput masne kiseline za energiju. Neuroni, međutim, zahtijeva stalni dotok glukoze, koja je u kratkom roku, pruža iz rezerve glikogena.
Kada je nivo insulina u jesen krvi, sintezu glikogena u smanjuje, jetre i enzime odgovorne za razgradnju glikogena postaju aktivni. Glikogen slom se stimuliše ne samo absense insulina, ali prisustvom glukagon, koji se luči kada je nivo glukoze u krvi padne ispod normalnih vrijednosti.
Insulin i metabolizam lipida
Metabolizma za korištenje masti i ugljikohidrata su duboko i neraskidivo isprepletene. duboke efekte s obzirom insulina na metabolizam ugljenih hidrata, logično je da inzulin također ima važne efekte na lipidni metabolizam, uključujući sljedeće:
1. Inzulin promovira sintezu masnih kiselina u jetri. Kao što je već rečeno, insulin je stimulativnog sinteze glikogena u jetri. Međutim, kao glikogen akumulira na visokom nivou (oko 5% mase jetre), dalje sinteza je snažno potisnut.
Kada je jetra zasićen glikogena, bilo kakve dodatne glukoze zauzima hepatocita je gurnuti u puteve koji vode do sinteze masnih kiselina, koje se izvoze iz jetre kao lipoproteini. U lipoproteini su pokidani u prometu, pružanje besplatne masne kiseline za upotrebu u drugim tkivima, uključujući adipocita, koji ih koriste za sintezu triglicerida.
2. Inzulin inhibira razgradnju masti u masnom tkivu inhibirajući intracelularne lipaze koji hidrolizira trigliceride za oslobađanje masnih kiselina.
Insulin olakšava ulazak glukoze u adipocitima, a unutar tih ćelija, glukoze može se koristiti za sintezu glicerola. Ovo glicerol, uz masne kiseline isporučene iz jetre, koriste se za sintezu triglicerida u adipocit. Do ovih mehanizama, inzulin je uključen u daljnje nakupljanje triglicerida u masnim ćelijama.
Iz cjelini perspektive tijela, inzulin ima učinak masti štede. Ne samo da vozi većina ćelija u preferencijalno oksidiraju ugljikohidrata umjesto masnih kiselina za energiju, inzulin posredno stimulira nakupljanje masti u masno tkivo.
Ostale značajne Efekti Inzulin
Osim inzulina efekt na ulazak glukoze u ćelije, i podstiče unos aminokiselina, ponovo doprinosi njenom ukupnom anabolički efekt. Kada je nivo insulina su niske, kao u postu državi, ravnoteža se gura prema intracelularni proteina degradacije.
Inzulin također povećava permiability mnogih ćelija kalij, magnezij i fosfat iona. Efekat na kalijum je klinički važno. Inzulin aktivira natrij-kalij ATPases u mnogim ćelijama, što je dovelo do tok kalija u ćelije. Pod određenim okolnostima, injekcije inzulina može ubiti pacijenata zbog svoje sposobnosti da akutno suzbijanja koncentracija u plazmi kalijuma.
Insulin Manjak i višak bolesti
Diabetes mellitus, nedvojbeno najvažniji metabolička bolest čovjeka, je nedostatak inzulina države. Takođe je značajan uzrok bolesti kod pasa i mačaka. Dva glavna oblika ove bolesti su prepoznati:
Tip I ili inzulin-neovisnim dijabetesom je rezultat frank nedostatka inzulina. Početka ove bolesti je obično u djetinjstvu. To je zbog uništenja beta ćelija pankreasa, najvjerojatnije rezultat autoimunosti na jednu ili više komponenti te ćelije. Mnogi od akutne efekte ove bolesti se može upravljati inzulina zamjene terapije. Održavanje stroga kontrola koncentracije glukoze u krvi monitoringom, insulin i dijetetskih upravljanje će smanjiti dugoročne negativne efekte ovog poremećaja na krvne sudove, nerve i drugih organskih sistema, što omogućava zdrav život.
Tip II ili ne-inzulin-neovisnim dijabetesom počinje kao sindrom inzulinske rezistencije. To je, ciljna tkiva ne da na odgovarajući način odgovori na inzulin. Tipično, početak ove bolesti je u odrasloj dobi. Uprkos monumentalna istraživačke napore, precizne prirode nedostataka koji vodi do dijabetesa tipa II je bilo teško utvrditi, i patogenezi ovog stanja je očito multifaktorijalan. Gojaznost je jasno jedan od glavnih faktora rizika, ali u nekim slučajevima ekstremne gojaznosti kod ljudi i životinja, osetljivost na insulin je normalno. Jer ne postoji, barem u početku, nemogućnost da se luče dovoljne količine inzulina, inzulin injekcije nisu korisni za terapiju. Umjesto toga bolest je pod kontrolom putem ishrane terapiju i hipoglikemici.
Hiperinsulinemija ili pretjerana lučenje inzulina je najčešće posljedica inzulinske rezistencije, povezani sa dijabetesom tipa 2 ili metabolički sindrom. Više rijetko, rezultati hiperinsulinemija iz insulina sekretujući tumora (insulinoma) u pankreasu. Hiperinsulinemija zbog slučajnog ili namjerno ubrizgavanjem prekomjerne inzulina je opasno i može biti akutno opasne po život zbog krvi glukoze pada brzo i mozak postaje gladi za energiju (inzulin šok).
Štitnjače hormonske receptore i Mehanizam djelovanja
Receptore za hormone štitnjače su intracelularni DNA-vezujući proteini koji funkcionišu kao hormon koji reaguje na faktore transkripcije, vrlo sličan koncepcijski na receptore za steroidne hormone.
Hormoni štitnjače ući ćelije kroz membranu transporter proteina. Jedan broj plazma membrane prevoznika su identificirani, od kojih su neki zahtijevaju ATP hidrolize; relativna važnost različitih nosača još uvijek nije jasno i mogu se razlikovati među tkiva. Jednom unutar jedra, hormon vezuje njegovih receptora, a kompleks hormon-receptor interakciju sa specifičnim sekvencama DNK u promotera odgovornih gena. Efekat kompleksa hormona receptora za DNK je da modulira ekspresiju gena, bilo poticanjem ili inhibiraju transkripciju specifičnih gena.
Za ilustraciju, razmotrite jedan mehanizam kojim tiroidnih hormona povećava snagu kontrakcije srca. Kontraktilnost srca ovisi, u dijelu, na relativni odnos različitih vrsta miozin proteina u srčani mišić. Transkripcija pojedinih miozin gena stimuliše hormone štitnjače, dok transkripcija drugih u inhibiran. Neto efekt je da se mijenja odnos prema povećan kontraktilnost.
Za dodatne informacije o mehanizmu djelovanja i kako su ti receptori u interakciji sa drugim faktorima transkripcije, ispitati sekciji hormona štitnjače receptori.
Fiziološki efekti hormona štitnjače
To je vjerojatno da sve ćelije u organizmu su mete za hormone štitnjače. Iako nije strogo neophodno za život, tiroidni hormoni imaju dubok uticaj na mnoge “Big Time” fiziološki procesi, kao što su razvoj, rast i metabolizam, a nedostatak hormona štitnjače nije kompatibilan sa normalnim zdravlje. Osim toga, mnogi od efekata hormona štitnjače su omeđeni studija deficita i višak država, kao što je nakratko ispod.
Metabolizam: hormoni štitnjače stimulirati raznovrsne metaboličke aktivnosti većini tkiva, što je dovelo do povećanja bazalnog metabolizma. Jedna od posljedica ove aktivnosti je da se poveća proizvodnja toplinu tijela, što se čini da će rezultirati, barem u dijelu, od povećane potrošnje kisika i stope ATP hidrolize. Putem analogije, akcija tiroidnih hormona je slično duva na tinjanje vatre. Nekoliko primjera specifičnih metaboličkih efekata hormona štitnjače uključuju:
Metabolizam lipida: Povećanje štitnjače hormona stimulirati masti mobilizacija, što dovodi do povećane koncentracije masnih kiselina u plazmi. Oni će povećati oksidaciju masnih kiselina u mnogim tkivima. Na kraju, koncentracije kolesterola i triglicerida su obrnuto proporcionalna sa nivoom hormona štitnjače – jedan dijagnostički indikta hipotireoze je povećana koncentracija holesterola u krvi.
Metabolizma ugljenih hidrata: tiroidni hormoni stimulišu gotovo sve aspekte metabolizma ugljikohidrata, uključujući poboljšanje insulin zavisni ulazak glukoze u ćelije i povećanje glukoneogeneze i glikokogenolizi da stvaraju slobodne glukoze.
Rasta: hormoni štitnjače su očigledno neophodne za normalan rast kod djece i mladih životinja, o čemu svjedoči i rast retardacije posmatrati u nedostatak štitnjače. Nije iznenađujuće, efekat promovira rast hormona štitnjače, usko je povezano sa onom hormona rasta, što je jasan indikta taj kompleks fizioloških procesa poput rasta ovise više kontrole endokrini.
Razvoj: Klasičan eksperiment u endokrinologija je bio dokaz da punoglavci lišena hormona štitnjače nije da se podvrgne metamorfoze u žabe. Od ključne važnosti u sisara je činjenica da normalan nivo hormona štitnjače su neophodni za razvoj fetusa i neonatalne mozga.
Ostali efekti: Kako je gore navedeno, čini se da ne bude organa i tkiva koji nisu pogođeni tiroidnih hormona. A nekoliko dodatnih, dobro dokumentirani efektima hormona štitnjače uključuju:
Kardiovaskularni sistem: hormoni štitnjače povećava broj otkucaja srca, kontraktilnost srca i minutni volumen. Oni također promovirati vazodilataciju, što dovodi do pojačane protok krvi na mnoge organe.
Centralni nervni sistem: Oba smanjena i povećane koncentracije hormona štitnjače dovodi do promene u mentalnom stanju. Premalo hormona štitnjače, a pojedinac ima tendenciju da se osjećaju mentalno sporog, dok previše izaziva anksioznost i nervozu.
Reproduktivni sistem: Normalno reproduktivnog ponašanja i fiziologija zavisi da uglavnom normalan nivo hormona štitnjače. Hipotireoza posebno je često povezana s neplodnošću.
Štitnjače bolesti države
Bolest je povezana i sa neadekvatne proizvodnje i hiperprodukcije tiroidnih hormona. Obje vrste bolesti su relativno česti nevoljama čovjeka i životinja.
Hipotireoze je rezultat od bilo koje stanje koje dovodi do nedostatka hormona štitnjače. Dva poznata primjera uključuju:
Nedostatka joda: jodid je apsolutno neophodan za proizvodnju hormona štitnjače; bez adekvatan unos joda, tiroidni hormoni ne može biti sintetiziran. Istorijski gledano, ovaj problem posebno je viđen u područjima s jodom-deficitarnim zemljišta, a Frank nedostatak joda je praktično eliminirana joda suplementacija soli.
Primarni štitnjače bolest: upalnih bolesti štitnjače koji uništavaju dijelovi žlijezde su jasno važan uzrok hipotireoze.
Uobičajeni simptomi hipotireoze nastao nakon ranog djetinjstva uključuju letargija, umor, hladno netoleranciju, slabost, gubitak kose i reproduktivnog neuspjeha. Ako ovi znaci su teške, kliničko stanje se naziva myxedema. U slučaju jodida nedostatak, štitaste postaje inordinantly velika i zove se gušavost.
Najteži i devestating oblik hipotireoze se vidi kod male djece sa nedostatkom urođenim štitnjače. Ako se takvo stanje ne ispraviti dopunskim terapije ubrzo nakon rođenja, dijete će patiti od kretenizam, oblik nepovratan rasta i mentalna retardacija.
Većina slučajeva hipotireoze se lako tretira oralne primjene sintetičkih hormona štitnjače. U prošlim vremenima, potrošnja Isušeno životinja štitne žlijezde je koristi za istu svrhu.
Hipertireoidizam rezultati lučenje hormona štitnjače. U većini vrsta, ovo stanje je manje uobičajena od hipotireoze. Kod ljudi najčešći oblik hipertireoze je Graves bolesti, imuni bolesti koje autoantitijela vezuju i aktivirati tireostimulišući receptora hormona, što dovodi do kontinuiranog stimulaciju sinteze hormona štitnjače. Još jedan zanimljiv, ali rijetki uzrok hipertireoze je tzv hamburger tireotoksikoza.
Uobičajeni znaci hipertireoze su u osnovi suprotna onima viđenim u hipotireoza, a uključuju nervoza, nesanica, visoka stopa srca, očne bolesti i anksioznosti. Graves bolest se obično tretiraju s anti-štitnjače lijekova (npr propylthiourea, methimazole), koji potiskuju sintezu hormona štitnjače prvenstveno ometa iodination od tireoglobulin od štitnjače peroksidaze.
Najpoznatiji komponenta želudačnog soka je solne kiseline, sekretornu proizvod parijetalni ili oxyntic ćelije. Poznato je da je kapacitet želuca da luči HCl je gotovo linearno u vezi sa brojevima parijetalne ćelije.
Kada je stimulirana, parijetalni ćelije luče HCl u koncentraciji od oko 160 mm (ekvivalent pH 0,8). Kiselina se luči u velike cannaliculi, duboko invaginacija plazma membrane koji su kontinuirano s lumen želuca.
Kada se stimuliše sekreciju kiselina je dramatične promjene u morfologiji membrane parijetalnog ćelije. Citoplazmatske tubulovesicular membrane koje su u izobilju u mirovanju ćeliji praktično nestati zajedno sa velikim povećanjem cannalicular membrane. Čini se da je protonske pumpe, kao i kalij i klorid provodljivosti kanala u početku nalaze na intracelularne membrane i prebačen u i spojene u cannalicular membranu neposredno prije sekreciju kiseline.
Na epitel želuca je suštinski otporna na štetne efekte želučane kiseline i druge uvrede. Bez obzira na to, prekomjerna lučenja želučane kiseline je glavni problem u ljudskim i, u manjoj mjeri, životinjske populacije, što dovodi do gastritisa, čira na želucu i bolesti peptičkog kiseline. Kao posljedica toga, parijetalni ćelija i mehanizme koristi da luči kiselina intenzivno su proučavali, što je dovelo do razvoja nekoliko lijekova korisnih za suzbijanje sekreciju kiseline.
Mehanizam sekreciju kiseline
Koncentracije vodikovih iona u parijetalni ćelija izlučevinama je oko 3 milijuna puta veća nego u krvi, a klorid se luči protiv i koncentraciju i električni gradijent. Prema tome, sposobnost partietal ćelije da luči kiselina zavisi od aktivnog transporta.
Ključni igrač u sekreciju kiseline je H +/K + ATPaze ili “protonske pumpe” koji se nalazi u cannalicular membrane. Ovo ATPaze je magnezijum-ovisna, a ne inhibitable po ouabain. Sadašnji model za objašnjavanje sekreciju kiseline je kako slijedi:
Jona vodonika se generišu unutar parijetalni ćelije od disocijacije vode. Hidroksilne ione formirana u tom procesu brzo u kombinaciji sa ugljen-dioksida za formiranje bikarbonata ion, reakcija cataylzed strane karboanhidraze.
Bikarbonata se transportuje iz basolateral membrane u zamjenu za hlorida. Odliv bikarbonata u rezultatima krvi u blagom uzvišenju pH krvi poznat kao “alkalne plima”. Ovaj proces služi za održavanje intracelularne pH u parijetalni ćelija.
Klorida i kalij iona se prenose u lumen cannaliculus strane provodljivost kanala, a kao što je potrebno za lučenja kiseline.
Jona vodonika se ispumpava iz ćelije, u lumen, u zamjenu za kalij kroz akciju protonske pumpe; kalij se na taj način učinkovito reciklira.
Akumulacija osmotski aktivni vodikov je ion u cannaliculus stvara osmotski gradijent kroz membranu koja rezultira spolja difuziju vode – rezultirajući sok želuca je 155 mm HCl i 15 mm KCl sa malom količinom NaCl.
Ključni supstrat za proizvodnju želučane kiseline je CO2, i širenje CO2 kroz bazalne površine parijetalnog čini se da je korak stopa ograničava u sintezi kiseline. Zanimljivo, ovo biohemijske princip je potvrđena proučavanjem želuca funkciju u aligatora. Ovi reptili proizvesti ogromne količine želučane kiseline nakon uzimanja velikog trupa, izgleda izobilju kiselina da su važni za ubrzavanje probave kostiju. Aligatori imaju vaskularne shunt koji preusmjerava CO2 -rich venske krvi u želudac, a ne direktno u pluća, povećanje količine CO2, koja se širi u parijetalni ćelija, a time i povećanje sintezu kiseline.
Kontrola sekreciju kiseline
Parijetalni ćelija nose receptore za tri stimulatori kiseline sekrecije, odražavajući triumverate neuralnih, parakrinu i kontrole endokrini:
Acetilkolin (muskarinski receptor tip)
Gastrina
Histamina (H2 receptor tip)
Histamina od enterohromafin poput ćelije može biti primarni modulator, ali pojavljuje se veličina stimulusa rezultira iz kompleksnog aditiva ili multiplikativni interakcije signala svake vrste. Na primjer, malim količinama histamina pušten stalno iz mastocita u želučane sluznice slabo stimuliše sekreciju kiseline, i slično za nizak nivo gastrina ili acetilkolina. Međutim, kada su prisutni nizak nivo svakog, sekreciju kiseline snažno prisiljen. Pored toga, farmakološki antagonisti svakog od tih molekula može blokirati sekreciju kiseline.Učinak histamina o parijetalni ćelija za aktiviranje adenilat ciklazu, što dovodi do visine od intracelularne ciklične koncentracije AMP i aktiviranje protein kinaze A (PKA). Jedan od efekata aktivacije PKA je fosforilacije citoskeleta proteina uključenih u transportu H + /K + ATPaze od citoplazme plazme membrane. Vezivanje acetilkolin i gastrin i rezultat u visini od intracelularne koncentracije kalcija.
Nekoliko dodatnih medijatora je dokazano da dovede do sekrecije želučane kiseline kada prožet u životinja i ljudi, uključujući kalcijum, enkefalina i bombezin. Kalcij i bombezin kako simuliraju gastrina oslobađanje, dok opijata receptori su identificirani na parijetalni ćelija. Nejasno je da li ove molekule imaju značajnu fiziološku ulogu u funkciji parijetalni ćelija.
Razne supstance su sposobni za smanjenje želuca lučenja kiseline kada prožet intravenski, uključujući i prostaglandina E2 i nekoliko peptida hormona, uključujući Sekretinska, želuca inhibitorni peptid, glukagon i somatostatin. PGE2, sekretinska i somatostatin može biti fiziološki regulatora. Somatostatin inhibira lučenje gastrina i histamina, a čini se da imaju direktan inhibitorni efekat na parijetalni ćelija.
Oksitocin u kiseline peptida devet amino koji je sintetiziran u hipotalamusa neurona i prevezli niz aksona zadnjeg hipofize za lučenje u krvi. Oksitocin se luči u mozgu i iz nekoliko drugih tkiva, uključujući i jajnika i testisa. Oksitocin se razlikuje od antidiuretskog hormona u dva od devet aminokiselina. Oba hormona se pakuju u granule i izlučuje zajedno s nosača proteina pod nazivom neurophysins.
Fiziološki efekti oksitocina
Proteklih godina, oksitocin je imao reputaciju kao “jednostavan” hormon, uz samo nekoliko dobro definisane aktivnosti koje se odnose na rođenje i dojenja. Kao što je bio slučaj sa toliko hormona, daljnja istraživanja pokazala mnogo suptilnih ali duboko uticaja ovog malog peptida, posebno u odnosu na njegove efekte u mozgu. Oksitocin je umiješan u postavljanje broj socijalnih ponašanja u vrsta u rasponu od miševa do ljudi. Na primjer, lučenje ili administraciji oksitocina kod ljudi pojavljuje za poboljšanje povjerenja i saradnje u okviru društveno-blizu grupe, uz promovisanje defanzivni agresija prema nepovezane, konkurentskih grupa.
Oksitocin je najbolje studirao u žena, gdje se jasno posreduje tri glavna efekta:
Stimulacija mlijeko izbacivanje (mlijeko mana): Mlijeko je u početku luči u male vrećice u mliječne žlijezde zove alveola, iz koje mora biti izbačena za potrošnju ili žetve. Mliječne alveole su okruženi glatkih mišića (mioepitelnih) ćelije koje su istaknutom meta ćelija za oksitocin. Oksitocin stimulira kontrakcije mioepitelnih ćelija, zbog čega mlijeko da se izbaci u kanale i cisterni.
Stimulacija materice kontrakcije glatkih mišića prilikom rođenja: Na kraju gestacije, materice mora energično i na duži vremenski period ugovora, kako bi se isporučiti fetus. Tokom kasnijim fazama trudnoće, postoji porast u izobilju oksitocina receptora na uterusa glatkih mišićnih ćelija, koja je povezana sa povećanom “razdražljivost” materice (a ponekad i majka, kao i). Oksitocin se oslobađa tokom porođaja, kada je fetus stimuliše cerviksa i vagine, a to povećava kontrakcije maternice glatkih mišića kako bi se olakšalo porod ili rođenja.U slučajevima kada kontrakcije materice nisu dovoljni za završetak isporuke, ljekari i veterinari ponekad upravlja oksitocin (“Pitocin”) da dodatno stimulisati kontrakcije materice – velika pažnja mora se obavljati u takvim situacijama kako bi se osiguralo da se fetus zaista mogu biti isporučeni i da izbjegavaju rupture materice.
Osnivanje majke ponašanja: Uspješno reprodukciju u sisara traži da majke postaju prilogu i odmah njeguju njihovi potomci nakon rođenja. Također je važno da se ne-dojilje žene ne manifestuju takve negovanje ponašanje. Istim događajima koji utječu na maternice i mliječne žlijezde u trenutku rođenja takođe utiču na mozak. U toku porođaja, došlo je do povećanja koncentracije oksitocina u likvora, a oksitocin glume u mozgu igra glavnu ulogu u uspostavljanju majke ponašanje.Dokaz za ovu ulogu oksitocina dolaze iz dvije vrste eksperimenata. Prvo, infuzija oksitocina u komore mozga Virgin štakora ili ne-trudna ovce brzo izaziva majke ponašanje. Drugo, administracija u mozak antitijela koja neutraliziraju oksitocin ili oksitocina antagonista će spriječiti majka štakora prihvati svoje mladunce. Druge studije podržavaju tvrdnju da je ova ponašanja učinak oksitocina je široko primjenjivo među sisarima.
Dok su svi efekata gore opisani sigurno javljaju kao odgovor na oksitocina, sumnja je nedavno bacila na nužnost u porođaja i majke ponašanje. Miševi koji nisu u stanju da luči oksitocin zbog ciljane poremećaja u oksitocina gena će se pare, dostave svoje mladunce bez očiglednog poteškoća i prikazuju normalno ponašanje majke. Međutim, oni pokazuju deficite u izbacivanje mlijeka i imaju suptilne poremećaj u ponašanju. To može biti najbolje da gledaju oksitocin kao glavni facilitator porođaja i majke ponašanje nego neophodna komponenta tih procesa.
Oba spola luče oksitocin – šta je njegova uloga u muškaraca? Muškarci sintetizirati oksitocina u istom područjima hipotalamusa kao kod žena, ali i unutar testisa, a možda i druge reproduktivne tkiva. Impulsi oksitocina se može otkriti prilikom ejakulacije. Trenutni dokazi pokazuju da oksitocin je uključen u olakšavanju sperme transport unutar muškog reproduktivnog sistema, a možda iu žena, zbog svoje prisustvo u sjemene tekućine. To može imati efekte na nekim aspektima muškog seksualnog ponašanja.
Kontrola oksitocina sekreta
Najvažniji stimulans za oslobađanje hipotalamusa oksitocina pokreće se fizička stimulacija bradavice ili sise. Čin dojenje ili sisanja se prenose u roku od nekoliko milisekundi do mozga preko kičmene refleks luk. Ovi signali utiču na neurone oksitocin-luči, što dovodi do oslobađanja oksitocina.
Ako želite dobiti ništa drugo osim trivijalnih količine mlijeka od životinja poput mliječnih krava, morate stimuliše oksitocin izdanju, jer je nešto poput 80% mlijeka dostupna samo nakon izbacivanja, i izbacivanje mlijeka zahtijeva oksitocin. Gledati neko pomuze kravu, čak i uz kavu, a ono što ćete vidjeti je da je prije muže, sise i donji vimena se pere nježno – ovo taktilna stimulacija dovodi do oksitocin puštanje na slobodu i izbacivanje mlijeka.
Veliki broj faktora može inhibiraju oksitocin oslobađanje, među njima akutnog stresa. Na primjer, oksitocin neuroni su potisnuti od strane kateholamina, koji se oslobađaju iz nadbubrežne žlijezde kao odgovor na mnoge vrste stresa, uključujući i strah. Kao praktični endokrini vrh – ne nose gorila kostim u mužu punu krava ili krenuli petardi oko majka doji svoju bebu.
Oba proizvodnju oksitocina i odgovor na oksitocin su modulirani strane kruže nivoa polnih steroida. Pucanje oksitocina pušten na rođenju izgleda biti izazvan dijelom cervikalni i vaginalni stimulacije fetusa, ali i zbog naglo pada koncentracije progesterona. Još jedan dobro proučenih učinak steroidnih hormona je značajno povećanje u sintezi uterusa (myometrial) oksitocin receptora krajem trudnoće, što je rezultat povećanja koncentracije cirkulirajućih estrogena.
Pripadnici roda Plasmodium su eukariotske mikrobe. Stoga, ćelija i molekularna biologija Plasmodium će biti sličan drugim eukariota. Jedinstvena karakteristika malarijskim parazita je njegova intracelularni način života. Zbog svoje intracelularne lokaciju parazit ima intimni odnos sa svojom ćelije domaćina koji se može opisati na ćelijskom i molekularnom nivou. Konkretno, parazit mora ući u ćelije domaćina, a jednom unutra, to mijenja ćelije domaćina. Molekularne i celularne biologije domaćina parazita interakcije su uključeni u ova dva procesa će se raspravljati.
Sadržaj:
Host eritrocita Invasion
Inicijalna Knjigoveške i MSP-1
Preorijentacija i Sekretorna Organelles
Junction Formiranje i Microneme Proteini
parazit Entry
Glisiranja motiliteta i Glidosome
Sažetak
Host eritrocita Datum
Kvake i Cytoadherence
Endotelne Cell Receptori
Antigeni Variation
Sažetak
Reference
Linkovi
Ćelije domaćina Invazija
Malarije paraziti su članovi Apicomplexa. Apicomplexa odlikuje skup organela u nekim fazama životnog ciklusa parazita. Ove organele, kolektivno poznate kao apikalni organele zbog lokalizacije na jednom kraju parazita, uključeni su u interakcije između parazita i domaćina. Konkretno, apikalni organele su uključeni u proces ćelije domaćina invazije. U slučaju Plasmodium, tri različita invazivnih oblika su identificirani: sporozoite, merozoite i ookinete (vidi Plasmodium Life Cycle). Sljedeće rasprava fokusira na ćelijskom biologije merozoites i eritrocita invazije. Reference na druge Apicomplexa i Plasmodium sporozoites će biti za ilustraciju zajedničke osobine.
Merozoites brzo (oko 20 sekundi) i posebno ući eritrocita.Ova specifičnost se ogleda i za eritrocita kao preferirani tip ćelije domaćina i za određenu vrstu domaćina, implicirajući receptor-ligand interakcije.Eritrocita invazija je složen proces koji se samo djelomično shvatiti na molekularnom i ćelijskom nivou. Međutim, značajan napredak je postignut u identifikaciji mnogih parazita i domaćina proteina koji su važni za proces invazije.
Proteini Merozoite površina i Host-Parazit Interakcije
Početne interakcije između merozoite i eritrocita je vjerovatno slučajni sudar i po svoj prilici uključuje reverzibilni interakcija između proteina na površini merozoite i domaćina eritrocita.Nekoliko površina proteina merozoite su opisani. Najbolji karakterizira je merozoite površinski protein-1 (MSP-1). Posredni o umiješanosti MSP-1 u eritrocita invazije uključuje njena ujednačena distribucija preko površine merozoite i zapažanje da antitijela protiv MSP-1 inhibira invaziju (Holder 1994). Osim toga, MSP-1does vezuju za band 3 (Goel 2003) I glycophorin A. Međutim, ulogu za MSP-1 u invaziji nije definitivno pokazao.Slično tome, circumsporozoite protein (SPRS) vjerojatno igra ulogu u ciljanje sporozoites na hepatocita kroz interakciju s heparinom sulfat proteoglikana (Sinnis i Sim 1997).
Još jedan zanimljiv aspekt MSP-1 je obrada proteolitičke da poklapa sa merozoite sazrijevanje i invazije (Cooper 1993). A primarne prerade javlja u vrijeme merozite sazrijevanja i rezultira u formiranju nekoliko polipeptida drže zajedno u ne-kovalentnim kompleksa. A sekundarne obrade javlja podudara sa merozoite invaziju na lokaciji u blizini C-terminus. Ne-kovalentnih kompleks MSP-1 polipeptid fragmenti je prolivena od površine merozoite sljedeće proteolize a samo mali C-terminalni fragment se vrši u eritrocita. Ovaj gubitak MSP-1 kompleks može korelaciji sa gubitkom ‘fuzzy’ kaput tokom merozoite invazije. C-terminalni fragment je pričvršćen na površinu merozoite strane GPI sidro i sastoji se od dva modula EGF-poput. EGF-poput modula mogu se naći u raznim proteina i obično se upleteni u protein-protein interakcije. Jedna mogućnost je da je sekundarna funkcija proteolitičke obrade izložiti EGF-poput modula koji jačaju interakcije između merozoite i eritrocita. Važnost MSP-1 i njegova obrada se podrazumijevaju iz sledeća zapažanja:
vakcinacija sa modulima EGF-kao da štiti od malarije i
Tačnu ulogu (e) koji nisu poznati MSP-1 i njegova obrada igra u procesu invazije merozoite. Ostali merozoite površina proteini također su uključeni u interation na merozoite sa eritrocita (komentar: Cowman 2012).
Preorijentacija i Sekretorna Organelles
Apikalni Organelles od
Plasmodium Merozoites
organele
oblik
Veličina (nm)
Microneme
ellipsoidal
40 x 100
Rhoptry
suza
300 x 600
gusta Granule
sferični
120-140
Nakon vezivanja eritrocita, parazit se okreće sama, tako da je “apikalni kraj” parazita je uporedo sa membrane eritrocita. Ovo merozoite preorijentacija se poklapa i sa prolaznim eritrocita deformacije. Apikalni membrana antigen-1 (AMA-1) je upleten u ovom preorijentaciju (Mitchell 2004). AMA-1 je transmembranski protein lokalizovan na apikalni kraju merozoite i vezuje eritrocite. Antitijela protiv AMA-1 ne mješati sa početnim kontakt između merozoite i eritrocita na taj način ukazuje na to da AMA-1 nije uključen u merozoite priključci. Ali antitijela protiv AMA-1 sprečavaju preorijentacija merozoite i time blokiraju merozoite invazije.
Specijalizirani sekretorni organele se nalaze na apikalni kraju invazivnih faza apicomplexan parazita. Tri morfološki različite apikalni organele detektuju elektronske mikroskopije: micronemes, rhoptries i guste granule (tabela). Guste granule nisu uvijek uključen u apeksnog organele i vjerojatno predstavljaju heterogenu populaciju sekretorni vezikule.
Sadržaj apeksnog organela su protjerani kao parazit napadne, sugerišući da su ove organele igraju neku ulogu u invaziji. Eksperimenti u Toxoplasma gondii ukazuju na to da je micronemes su protjerani prvi i doći sa inicijalni kontakt između parazita i domaćina (Carruthers i Sibley 1997). Povećanje citoplazmatske koncentracije kalcija i cAMP (Dawn 2014) je povezana sa microneme pražnjenja i može uključivati put signala uključuje fosfolipaze C, inositoltriphosphate i kalcijum zavisne protein kinaze (Sharma i Chitnis 2013).
U rhoptries se ispuštaju neposredno nakon micronemes i oslobađanje njihovog sadržaja javlja se u dva koraka koji uključuje prvo vrat rhoptry zatim sijalica u rhoptry.
Gusta granula sadržaji su pušteni nakon što je parazit završi svoj ulazak, i stoga su obično uključeni u modifikaciju ćelije domaćina. Međutim, subtilisin nalik proteaze, koji su upleteni u sekundarnoj obradi proteolitičke MSP-1 (gore navedeno), takođe su lokalizirane na Plasmodium guste granule (Blackman 1998, Barale 1999). Ako je MSP-1 za obradu katalizovane ovim proteaze, onda barem neke guste granule moraju biti otpušteni u vrijeme invazije.
Specifičnih interakcija i Junction stvaranja
Sljedeći merozoite preorijentacija na micronemes obavljati njihov sadržaj. Ove microneme proteini sadrže mnogo proteina koji se zna da su adhesins i obavezujuće ovih adhesins za receptore na domaćina eritrocita jača interakcija između eritrocita. Proteini lokaliziran na micromenes uključuju:
EBA-175, a 175 kDa ‘eritrocita vezivanje antigena’ iz P. falciparum
DBP, Duffy-vezujući protein iz P. vivax i P. knowlesi
SSP2, Plasmodium sporozoite površinski protein-2. Također poznat kao TRAP (protein ljepilo thrombospondin u vezi).
Proteini sa homologije na SSP2/TRAP iz Toxoplasma (MIC2), Eimeria (Etp100), i Cryptosporidium
CTRP, circumsporozoite- i TRAP-u vezi proteina Plasmodium nalaze u fazi ookinete
Receptora / liganda Interakcije
vrsta
Host receptora
Merozoite liganda
P. falciparum
glycophorins (sijalične kiselina)
EBA-175
P. vivax,
P. knowlesi
Duffy antigen
DBP
Od posebnog značaja su EBA-175 i DBP koji prepoznaju ostataka sijalične kiselina glycophorins i Duffy antigen, (Tabela). Drugim riječima, ovi parazit proteini su vjerojatno uključeni u receptor-ligand interakcije sa proteinima izložene na površini eritrocita. Poremećaj rezultata EBA-175 gena u parazit prebacivanje iz sijalične kiseline zavisi od puta do sijalične kiseline nezavisan put (Reed 2000), što ukazuje da postoji neka redundancije u odnosu na interakcije receptor-ligand. Zaista, nekoliko proteina koji se odnose na EBA-175 su identificirani u P. falciparum i čine gen porodice kao (EBL) proteini eritrocita vezivanja (Tham 2012).
Poređenje sekvenci EBA-175 i DBP otkrivaju konzervirani strukturne karakteristike koje su također dijeli s drugim EBL proteina. Među njima su transmembranski domene i receptora vezivanja domena (slika, modificirani iz Adams 1992). Aktivnost vezivanja za receptore je mapiran na domenu u kojoj su konzervirani cistein i ostataka aromatskih aminokiselina između vrsta (plave područje na slici). Ovaj navodni obavezujući domena duplicirane EBA-175. Topografija transmembranski domena je u skladu sa ligandi parazit kao integralni membranski proteini sa domenom receptora vezivanja izložene na površini merozoite sljedeće microneme pražnjenja.
Još jedna porodica adhesins uključenih u vezivanju merozoites na erthrocytes su retikulocita vezivanja poput kolegama (RH).Članovi ove porodice udjela homologije s protein prvobitno identifikovan u P. vivax koji se vezuje specifično retikulocita i mogu igrati ulogu u retikulocita specifičnosti P. vivax. Ostali microneme proteina u ‘TRAP’ porodice su također uključeni u lokomotorni i/ili ćelija invaziju na pozornici sporozoite i drugih apicomplexa (Tomley i Soldati 2001). Svi ovi proteina (EBL, Rh, TRAP obitelji) imaju domene koje su navodno uključeni u ćeliji ćelija prijanjanje, kao i trans-membrana domene na svom C-termini. Microneme (Mn) otpuštanje će izlagati ljepilo domene, a potom bi ih vežu za receptore na ćelije domaćina i time čine vezu između invazivni oblik (npr merozoite ili sporozoite) i ćelije domaćina (Slika).
U slučaju merozoite invazije ove interakcije je posredovana između nekoliko članova EBL i Rh porodice. Ovi različiti adhesins vežu za različite receptore na eritrocita i pružiti redundantnost u merozoite vezivanje za eritrocita (Tham 2012). Jedan od elemenata ovog redundancije predviđa back up plan u slučaju jedne ligand/receptora par ne uspije. Na primjer, ako je u stanju da blokira njegove interakcije sa svojim receptor je odgovor antitijela protiv jednog od parazita liganada, onda postoje i drugi ligandi i receptori koji mogu ispuniti ovu ulogu u obavezujuća. Osim toga, uključivanje više parova ligand/receptora u funkciji istovremeno će ojačati interakciju između parazita i ćelije domaćina.
Ova interakcija je dodatno ojačana oslobađanje dva dodatna protein kompleksa iz regiona vrat rhoptries (Weiss 2016). Jedan od njih je kompleksan uključuje RH5 (slika). RH5 vezuje za eritrocita površinski protein poznat kao basigin. U basigin receptora može biti od ključnog značaja za invaziju P. falciparum (Crosnier 2011). RH5 je vezan za merozoite kroz interakciju sa RH5 interakciji proteina (Ripr) koji također veže za merozoite površinski protein poznat kao cystiene bogat zaštitni antigen (CyRPA). RH5 Vjeruje se da igra važnu ulogu u invaziji u tome da se štedi preko Plasmodium vrsta i očigledno ne mogu biti nokautiran. Ostalim članovima Rh porodice, kao i pojedinih članova EBL porodice, čini se da više dispensible.
Još jedan proteinski kompleks pušten iz vratu rhopthries uključuje grupu proteina poznat kao Röns za rhoptry vrat. U Röns se ubacuju u domaćina membranu nakon puštanja (Slika). Ron2 vezuje za AMA-1 koji je lokaliziran na površini merozoite (Tonkin 2011). U ovom slučaju parazit snabdeva i ligand i receptor. U ron2/AMA-1 kompleksa zatim doprinosi i ovu vezu formirana između merozoite i eritrocita i dalje jača veza između parazita i domaćina (Weiss 2016). Osim toga, ron2 i AMA-1 su visoko konzervirani acrossed je Apicomplexa ukazuje na centralnu ulogu ovih proteina u procesu invazije.
Poklapa sa lučenje ovih različitih liganada i njihovih interakcija sa različitim receptorima je pojava elektrona guste spoju između eritrocita i merozoite (Slika). Uske formiranje spoju može pokrenuti microneme pražnjenja slijedi oslobađanje rhoptry vrata proteina koji izlaže domene vezivanja za receptore parazita liganda. Po svoj prilici ovo čvrsto spoju se sastoji od ovih različitih receptora/ligand interakcije. Sa svakim uzastopnim oslobađanje ligand i njegova vezivanja za svoj receptor na lakomost od merozoite/eritrocita interakcije povećava.
Arrow označava elektrona guste spoj između merozoite i eritrocita. Mikrograf od Aikawa et al (1978) J. Cell Biol. 77:72.
Ukratko:
elektron-guste spoju forme između apikalni kraj merozoite i domaćin membrane eritrocita odmah nakon preorijentacija
čvrsto formiranje spoju se podudara sa oslobađanje micronemes i regiona vrat rhoptries
proteini oslobodili od parazita vezuju za receptore na površini eritrocita ostajući priključen na merozoite
posebno značajan ligand/receptora par su ron2 i AMA-1 koji su oba isporučuje parazita i igrati ključnu ulogu u formiranju uske spoju
Parazit Ulaz
Apicomplexan paraziti aktivno invaziju ćelija domaćina i stupanje nije zbog uzimanja ili fagocitozu od strane ćelije domaćina. Ovo posebno dolazi do izražaja u slučaju eritrocita koji nedostaje sposobnost fagocitne. Osim toga, membrane eritrocita ima 2-dimenzionalni submembrane citoskeletonom koji isključuje endocitoze. Stoga je poticaj za formiranje parasitophorous vakuole mora doći od parazita. Nekoliko događaja dogoditi prilikom ulaska parazita, uključujući: 1) poremećaja u submembrane citoskeleta od eritrocita, 2) formiranje parasitophorous vakuole, i 3) i prolijevanje merozoite površine proteina. Parazit unos prešao strane acto-miozin motor kompleks zove glidosome.
Membrane eritrocita proteini se redistribuira u trenutku formiranja spoju tako da kontaktna površina je besplatan eritrocita membranskih proteina. A merozoite serin proteaza koja cijepa eritrocita bend 3 je opisana (Braun-Breton 1993). Zbog ključnu ulogu bend 3 predstave u homeostatis u submembrane skeleta, njegove degradacije može dovesti do lokalizirane prekid citoskeleta. Reorganizacija submembrane citoskeleta i lipida arhitektura vjerojatno prati merozoite invazije (Zuccala 2011).
Jedan početne parasitophorous vakuolarnom membrana (PVM) formi u spoju području. Ova membrana invaginacija vjerovatno potiče iz oba domaćina membrane parazita komponente i širi kao parazit ulazi u eritrocita. Veze između rhoptries i nastajanju PVM se ponekad posmatra (slika, strelica). Osim toga, sadržaj rhoptries su često lamelarne (tj višeslojni) membrane, a neki rhoptry proteini su lokalizirane na PVM sljedeće invazije, što ukazuje da je rhoptries funkcioniraju u PVM formiranju (Sam-Yellowe 1996).
Mikrograf od Aikawa et al (1978) J. Cell Biol. 77:72
Ookinetes nedostaje rhoptries i ne formiraju parasitophorous vakuole unutar komarca midgut epitelne ćelije. U ookinetes brzo prolaze kroz epitelne ćelije i izazvati veliku štetu da krenu prema bazalne lamine (Han 2000, Ziegler 2000). Slično tome, sporozoites mogu ulaziti i izlaziti hepatocita bez prolazi exoerythrocytic schizogony. Oni paraziti koji ne prođu schizogony su slobodni u domaćinu citoplazmi, dok su oni u fazi schizogony su zatvorene unutar PVM (Mota 2001). Ova zapažanja ukazuju na to da je potrebna PVM za intracelularne razvoj i nije neophodan za proces ćelije domaćina invazije. Kao što se formira početni parasitophorous vakuole, spoju (označen sa C na slici) između parazita i domaćina postaje prsten nalik i pojavljuje se parazit za kretanje kroz ovaj anulusa jer ulazi u širi parasitophorous vakuole. Umjesto toga ovaj kreće spoju se izvukao iz ispred parazita do zadnjeg što je dovelo do Kretanje parazita u ćelije domaćina.
Kao parazit ulazi, MSP-1 mnogi od merozoite površine proteini se prolio. Ovaj proces rasterećenja posredovana specifične proteaze te je naredio proces (Boyle 2014).
The Glideosome
Model kreće spoju kompleksa i glidesome vožnje klizi motilitet iz Besteiro (2011).
Invazivnog oblika apicomplexan paraziti su često pokretnih oblika da je puzati duž supstrat jednom vrstom pokretljivost nazivaju ‘klizi pokretljivost. Klizeći pokretljivost, kao što je invazija, uključuje i oslobađanje adhesins, vezanost za supstrat, a prekrivanje je adhesins na zadnjem kraju zoite. Jedna razlika između zmajem pokretljivost i invazija je da micronemes i rhoptires moraju biti stalno objavljena kao organizam kreće.Tako je, klizeći pokretljivost ne uključuje ovaj relativno mali kreće spoju, ali kontinuirano stvaranje novih spojeva između zoite i supstrata. Osim toga, adhesins su cepane od površine zoite kao priraslica do zadnjeg od zoite i trag ljepila molekula ostavio iza kreće zoite na podlozi. Mehanizam pokretljivost i invazije su vrlo slični i na taj način, za vrijeme invazije parazita doslovno puzi u ćelije domaćina kroz raskrsnice kreće. Osim toga, neke apicomplexans koristiti ovaj tip pokretljivost pobjeći iz ćelija i može preći biološke barijere ulasku i izlasku iz ćelije.Kompleksa proteina koji pokreće ovo klizi motilitet je poznata kao glidosome (Boucher 2015).
Cytochalasins inhibiraju ulazak merozoite, ali ne i vezanosti, čime se ukazuje na to da je potrebno snage za parazita invazije i klizeći pokretljivost se temelji na aktin-miozin citoskeleta elemenata. Sposobnost miozin, a motor protein, za stvaranje sile je dobro poznata (npr., Mišićne kontrakcije). A miozin jedinstven za Apicomplexa je identificiran i usidren u unutrašnje membrane kompleks (IMC). IMC se odnosi na dvostruke membrane leže pod plazma membrane na invazivne fazama Apicomplexan parazita. Ovaj IMC je dodatno podržan od strane pod-sferične mikrotubule koje se protežu dužinom parazita. IMC povezana miozin interakciju sa aktin kao dio glidesome. Različiti adhesins (tj EBL, Rh, TRAP i AMA-1) koji čine kretanje spoju (MP) kompleks se zatim povezana sa glidesome (Slika).
Članovi porodice TRAP i drugih adhesins imaju konzerviranom citoplazmatski domena. Ovaj citoplazmatske domene je povezano sa kratkim aktin vlakna preko Aldolaza. Na aktin vlakna i miozin su orijentirani u prostoru između unutrašnje membrane kompleks i plazma membrane, tako da miozin pokreće aktin vlakna prema zadnjem u zoite. Na miozin je usidren u IMC-a i ne miče. Stoga, transmembranski adhesins se izvukao kroz fluid lipida dvosloja plazma membrane zbog njihove povezanosti sa aktin vlakna. Tako je kompleks adhesins i aktina niti se prevozi prema zadnjem ćelije. S obzirom da je adhesins su ili kompleksni sa receptora na ćelije domaćina i usidren u ćeliju citoskeleton domaćina, ili vezani za supstrat, neto rezultat je naprijed kretanje parazita (slika). Kada je adhesins do kraja zadnjeg parazita su proteolyitcally Cepane i šupe iz zoite površine.
U slučaju ćelije invazije PVM i domaćin ćelijske membrane će morati da zapečaćeni, tako da je PVM je netaknut i okolnih parazita i domaćina plazma membrana je netaknuta. Mehanizmi su uključeni u ovaj zadnji korak invazije nisu poznati.
Identifikovani su mnoge proteine koji su uključeni u proces invazije. To će uključivati signalizacije između različitih koraka invazije (Santos i Soldati-Favre 2011). Međutim, mnogo toga ostaje da se saznali ćelijske i molekularne biologije merozoite invazije. A bolje razumijevanje složenog procesa parazita invazije moglo dovesti do razvoja novih terapijskih pristupa malarije i drugih bolesti uzrokovanih Apicomplexans.
Sažetak
Merozoite invazija je kompleksan i naredio proces. A okvirni model merozoite invazije uključuje:
Inicijalna merozoite vezivanje uključuje reverzibilni interakcija između površine proteina merozoite i domaćin erythrocyte.The točne uloge MSP1 i drugih merozoite površinski proteini nisu poznati.
Preorijentacija nepoznati rezultati mehanizam u apikalnoj kraju merozoite se uporedo sa membrane eritrocita.
Pražnjenja micronemes i vrat je rhoptries poklapa sa formiranjem uske spoj između domaćina i parazita.
Uske spoju posredovana receptorom-ligand interakcija između površine proteina eritrocita i Integral parazit membranski proteini izloženi ispuštanjem apeksnog organela.
Lokaliziran čišćenje eritrocita submembrane citoskeleta i formiranje početne parasitophorous vakuole (PVM) je u korelaciji sa kompletnom pražnjenja rhoptries.
Uske spoju postaje u obliku prstena i povukao prema nazad u merozoite da prisili merozoite u formiranju parasitophorous vakuole.
Sila nastaje miozin motora povezan sa trans-membrane parazita ligandi, koji se zove glidosome, kreće duž aktin vlakna unutar parazita.
Invazija je završen do zatvaranja PVM i membrane eritrocita.
HOST ERITROCITA IZMJENE
Jednom unutar eritrocita, parazit prolazi trofičkim faze slijedi replikativnu fazu. Tokom ovog intraerythrocytic perioda, parazit mijenja domaćin čine ga više prikladan staništa. Na primjer, membrane eritrocita postaje propušta male metabolita molekularne težine, pretpostavlja se odražava potrebe aktivno raste parazita je (vidi UptakeX i propusnosti).
Još jedna modifikacija ćelije domaćina tiče cytoadherence od P. falciparum – inficiranim eritrocita za endotelne stanice, a samim tim sekvestracija zrelog parazita u kapilare i post-kapilarna venula. Ovo sekvestracija vjerojatno dovodi do microcirculatory izmjene i metaboličkih poremećaja koji bi mogao biti odgovoran za mnoge od manifestacija teške falciparum malarije (vidi patogenezi). Je cytoadherence za endotelne stanice savetuje najmanje dvije prednosti za parazita: 1) mikroaerofilne okruženje koje je pogodnije za metabolizam parazita, i 2) izbjegavanje slezine i kasnije uništavanje.
Kvake i Cytoadherence
A velike strukturalne promene domaćina eritrocita su elektron-guste izbočina ili “dugmad”, na membrane eritrocita od P. falciparum – inficiranim ćelija. Dugmad su izazvane parazita i nekoliko parazit proteina su povezani sa dugmadi (Deitsch i Wellems 1996). Dva proteina koji mogu sudjelovati u formiranju dugme ili utjecati na domaćina eritrocita submembrane citoskeletonom i indirektno izazivaju formiranje dugme su ručku povezane histidin bogat proteinima (KAHRP) i eritrocita membranski protein-2 ( Pf EMP2), koji se nazivaju MESA. Ni KAHRP ni PfEMP2 su izloženi na vanjskoj površini eritrocita, ali su lokalizovana na citoplazmatski lice domaćina membrane (slika). Njihova tačna uloga u formiranju dugme nisu poznati, ali može uključivati reorganizaciju submembrane citoskeleta.
Dugmad se vjeruje da igraju ulogu u sekvestracija zaraženih eritrocita jer su točke kontakta između zaraženi eritrocita i vaskularnih endotelnih ćelija i vrsta parazita koji izražavaju gumbi pokazuju najviši nivo zaplene. Osim toga, prekid KAHRP rezultira gubitkom dugmad i sposobnost da cytoadhere pod uvjetima protoka (Crabb 1997). A polimorfne protein, nazvan Pf EMP1, takođe je lokaliziran na dugmad i izložen na površini domaćina eritrocita. Translokacija od Pf EMP1 na površinu eritrocita ovisi u dijelu na drugom membrane eritrocita povezan protein Pf EMP3 (Waterkeyn 2000). pfEMP1 vjerojatno funkcionira kao ligand koji se vezuje za receptore na matičnom endotelnih ćelija. Druge predložene cytoadherence ligandi uključuju modificirani band-3, pod nazivom pfalhesin (Sherman 1995), sequestrin, rifins i clag9 (Craig i Scherf 2001).
Pf EMP1 je član var gena porodice (Hviid 2015). Svaki parazit ima oko 40-60 var gena koji pokazuju visok stepen varijabilnosti, ali imaju sličnu ukupnu strukturu (slika). PfEMP1 ima veliki ekstracelularnog N-terminalni domena, a transmembranski regija i C-terminalni intracelularne domene.Regije C-terminalni se štedi između članova var porodice i vjeruje se usidri Pf EMP1 na eritrocita submembrane citoskeleta. Konkretno, ova kisela C-terminalni domene može komunicirati sa osnovnim KAHRP od knaufa (Waller 1999), kao i spektrin i aktin (O 2000).
Ekstracelularne domene odlikuje 1-5 kopije Duffy vezivanja kao što je (DBL) domena. Ove DBL domeni su slične regiji receptor vezivanje liganda uključenih u merozoite invazije (gore navedeno). U DBL domeni izložba konzerviranog razmakom od cisteina i hidrofobnih ostataka, ali inače pokazuju malo homologije. Filogenetski analiza pokazuje da postoji pet različitih klasa (označen kao jedan , b , g , d , i e) u DBL domena (Hviid 2015). Prvi DBL je uvijek isti tip (određen je), A to je praćeno InterDomain regija cistein-bogatih (CIDR). Varijabla broj DBL u raznim naloga čine ostatak ekstracelularne domena Pf EMP-1.
Tokom svakog mitotska ciklusa var gena podvrgnuti rekombinacije dovodi do kontinuiranog generacije dodatnih varijanti (Claessens 2014). Zanimljivo je da je struktura var gena porodica je prisutan još od prije P. falciparum i P. reichenowi divergencija koja se dogodila prije više od dva miliona godina (Zilversmit 2013).
Endotelne Cell Receptori
Mogući Receptori Utvrđene
In Vitro Analize vezivanja
*CD36
*ig Natporodica
ICAM1
VCAM1
PECAM1 Hondroitin sulfat A +endotelne protein C +receptor
+heparan sulfat
+hijaluronska kiselina
+E-selektin
+thrombospondin +Rosetting Ligandi
CR-1
krvne grupe A Ag
glikozaminoglikana
Nekoliko mogućih endotela receptora (Box) su identifikovane testiranjem sposobnost zaraženih eritrocita za vezivanje u statičkim pridržavanje testovima (Beeson and Brown 2002). Jedan od najboljih karakterizira među njima je CD36, što je 88 kDa integralni membranski protein koji se nalazi na monocite, trombocita i endotelnih ćelija. Zaražene eritrocita od većine parazit izolira se vezuju za CD36 i vezivanje domena je mapiran na CIDR od Pf EMP1 (vidi sliku). Međutim, CD36 nije otkriven na endotelne stanice moždane krvne sudove i parazita iz kliničkih izolata imaju tendenciju da se pridržavaju i CD36 i intracelularne adhezije molekula-1 (ICAM1). ICAM1 je član imunoglobulina natporodice i funkcije u ćeliji ćelija prianjanja. Osim toga, sekvestracija zaraženih eritrocita i ICAM1 izraz ko-lokaliziran u mozgu (Turner 1994).
Hondroitin sulfat A (CSA) je upleten u cytoadherence u placentu i može doprinijeti negativnih efekata od P. falciparum tokom trudnoće. Uloga nekih drugih potencijalnih receptora nije jasno. Na primjer, pridržavanje thrombospondin pokazuje nizak afinitet i ne može podržati obavezujuće pod uvjetima protoka. Vezivanja VCAM1, PECAM1 i E – selektina čini se da je rijetko i pitanja u vezi sa njihovim konstitutivnim izraz na endotelne stanice su podignute. Međutim, cytoadherence moglo uključivati više receptor/ligand interakcije.
Rosetting je još jedan ljepilo fenomen ispoljava P. falciparum – inficiranim eritrocita. Zaraženim eritrocitima iz nekih parazit izolata će vezati mutiple nezaraženih eritrocita i Pf pojavljuje EMP1 da ima ulogu u barem neke rosetting. Mogući receptori uključuju dopuna receptor-1 (CR1), krvna grupa A antigena, ili glikozaminoglikana polovinama neutvrđenog proteoglikana. (Vidi sliku koja prikazuje moguće receptor-ligand interakcija uključenih u rosetting na drugu web stranicu).
Različite vrste DBL domena i CIDR (gore navedeno) vezuju za različite endotelne ćelije receptore (Craig i Scherf 2001). Na primjer, DBL a, koji se sastoji od prve domene, vezuje za mnoge receptora povezanih s rosetting. Vezivanje CIDR na CD36 može objasniti obilje ovog konkretnog obavezujuće fenotip među parazit izolata.
Kodiranje cytoadherence ligand od strane visoko polimorfne gen porodice predstavlja paradoks u tom receptor/ligand interakcije se općenito smatra vrlo specifičan.Zanimljivo je izbor za različite cytoadherent fenotipova rezultirati concommitant promjenu u vrsti površine antigene (Biggs 1992). Slično tome, ispitivanje klonskih parazita linija otkrio da su promjene u vrsti površine antigenske korelaciji sa razlikama u vezivanja za CD36 i ICAM1. Na primjer, roditeljske linije (A4) poštuje jednako dobro za CD36 i ICAM1, dok je jedna od A4-izvedeni klonove u (C28) izloženi značajnog prednost za CD36 (slika, modificirani iz Roberts 1992). Vezivanje za ICAM1 je zatim ponovo izabran prelazite zaraženih eritrocita na ICAM1.Sva tri parazit klonovi (A4, C28, C28-I) izlagao različite antigene tipa kao što je pokazano aglutinacije sa hiper-imuni Sera.
Izraz određenog PfEMP1 će rezultirati parazit sa jasnim cytoadherent fenotip, a to može utjecati na patogenezu i bolesti ishod. Na primjer, vezivanje za ICAM-1 je obično umiješan u moždane patologije. Stoga, paraziti izražavajući PfEMP1 koji se veže za ICAM1 može biti više vjerojatno da će izazvati cerebralne malarije. U stvari, viši nivo transkripcije posebno var gena nalaze se u slučajevima teške malarije u odnosu na jednostavan malarije (Rottmann 2006). Slično tome, veći udio izolata koji se vezuju na ADU su dobijeni iz placente u odnosu na perifernu cirkulaciju bilo trudnice ili djeca (slika, modificirani iz Beeson 1999). Osim toga, posteljice malarija je često povezana sa višim nivoima transkripcije određenog var gen koji povezuje CSA (Duffy 2006). Ovaj fenomen nije ograničen samo na posteljicu u da postoji dominantan izraz određenog var gena u različitim tkivima (slika, od Montgomery 2007). Kasniji rad je potvrdio da različite varijante PfEMP1 imaju različite tropisms za različita tkiva (Smith 2014).
Slika iz Montgomery 2007. Pokazuje broj različitih tipova PfEMP1 (označen kao grupa 1-6) izražene u različitim tkivima (mozak, pluća, srce i slezina) iz 3 različite pacijenata. PM30 je umro od teške malarije anemije. PM32 je dijagnosticiran i cerebralne malarije i teške anemije. PM55 je dijagnosticiran samo cerebralne malarije.Figure, modificirani iz Beeson 1999 . Pokazuje proporcije izolata koji se vezuju za CSA, CD36, ili ICAM-1. Zaraženim eritrocitima su prikupljeni iz placente, periferna cirkulacija majke, ili periferne cirkulacije djeteta.
Još nedavno je dokazano da je poseban podskup var geni su vrlo prepisuju sljedeći izbor na ljudski mozak endotelu i da ti isti različita podtipa su povezani sa cerebralnom malarije (Aird 2014; Cunnington 2013; Smith 2013). Ovo tkivo specifični izraz određenog var gena ukazuje na to da različita tkiva biraju različite parazit populacije na osnovu konkretnih PfEMP1 se izrazio na površini zaraženih eritrocita.
Iako sekvestracija nudi mnoge prednosti parazita, izraz antigena na površini zaraženih eritrocita daje cilj za imuni sistem domaćina. Parazita kompenzuje imunološki odgovor domaćina izražavajući antigenically različite Pf EMP1 molekule na površini eritrocita. Ovo omogućava parazit izbjeći izbija imunog sistema domaćina, ali još uvijek održava cytoadherent fenotip. Ovaj antigeni prebacivanje može doći do onoliko često koliko je 2% po generaciji u odsustvu imunološkog pritiska (Roberts 1992). Molekularni mehanizam antigeni prebacivanje nije poznat.Eksperimentalni dokazi ukazuju da je mehanizam nije povezan sa dvostrukih transpoziciju u određene lokacije izraz povezanih kao naći u afričkim tripanozoma. Samo jedan var gen je izražen u vremenu (tj alelne isključenje). Ne-izrazio gena šutio od proteina koji se vezuju na promotor regije. A gen može doći uskoro izjavom da na određenoj lokaciji u jezgru i povezana je sa hromatina modifikacija. Ovaj izraz mjesto može primiti samo jednu aktivnu gena promotora. Tako je var promoter je dovoljan i za prigušivanje zvuka i mono-alelne transkripciju PfEMP1 alela (Voss 2006; Guizetti 2013).
Sažetak
Malarijskim parazit mijenja eritrocita izvozeći proteina u ćelije domaćina.
Jedan od takvih modifikacija je izraz Pf EMP1 na eritrocita površini koja funkcionira kao cytoadherent ligand.
Vezivanje ovog ligand za receptore na matičnom endotelu promovira sekvestracija i omogućava zaraženih eritrocita izbjeći slezine.
Brojni Pf EMP1 gena (tj var gen porodice) pružiti parazit sa sredstvima varirati antigen izražene na površini eritrocita.
Ovaj antigeni varijacija također korelira s različitim cytoadherent fenotipova.
Nervnog sistema vrši veliki uticaj na sve probavni procesi, naime pokretljivost, ion transport povezan sa lučenjem i apsorpciju, i gastrointestinalnog protok krvi. Neki od ove kontrole izvire iz veze između probavnog sistema i centralnog nervnog sistema, ali jednako važno, probavnog trakta je obdarena sa vlastitim, lokalnim nervni sistem naziva crijevne ili unutrašnje nervnog sistema. Veličine i složenosti crijevne nervnog sistema je ogroman – sadrži onoliko neurona kao i kičmene moždine.
Na crijevne nervni sistem, zajedno sa simpatički i parasimpatički nervni sistem, čine autonomni nervni sistem.
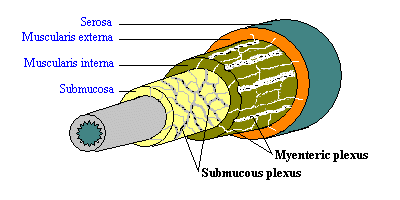
Glavne komponente crijevne nervnog sistema su dvije mreže ili pleksusa neurona, a oba su ugrađeni u zidu probavnog trakta i protežu od jednjaka do anusa:
Je mijenteričnom pleksus se nalazi između uzdužne i kružne slojeva mišića u tunike mišićni i, na odgovarajući način, vrši kontrolu prvenstveno preko probavnog motilitet trakta.
Je mukozna pleksus, kako mu i ime govori, je sahranjen u submukoze. Njegova glavna uloga je u sensing okruženje unutar lumena, kojim se uređuje gastrointestinalne protok krvi i kontrolu epitelnih funkciju ćelije. U područjima gdje su minimalne ove funkcije, kao što je jednjak, je mukozna pleksusa je oskudan i može zapravo nedostaje u rubrikama.
Slika ispod prikazuje dio mijenteričnom pleksusa u odjeljku mačke duodenuma. Pass pokazivač miša preko slike da bi navesti nekoliko crijevne neurona.
Pored dva glavna crijevne nervnih pleksusa, postoje manji pleksusa ispod serosa, u kružni glatkih mišića i sluznice.
Unutar crijevne pleksusa su tri vrste neurona, od kojih je većina multipolarnog:
Senzorni neuroni primaju informacije od senzornih receptora u sluznici i mišića. Najmanje pet različitih senzornih receptora su identifikovani u sluznici, koji odgovaraju na mehanički, termički, osmotski i kemijskih podražaja. Hemoreceptori osjetljive na kiseline, glukoze i aminokiselina su pokazali koji, u suštini, omogućuje “degustacija” od lumenal sadržaja. Senzornih receptora u mišićnim odgovor da se protežu i napetosti. Kolektivno, crijevne senzorne neurone sastaviti sveobuhvatan baterija informacija o Gut sadržaja i stanja gastrointestinalnog zida.
Motor neurona unutar crijevne pleksusa kontrolu gastrointestinalnog pokretljivost i lučenje, a možda i apsorpciju. U obavljanju ove funkcije, motorni neuroni djeluju direktno na velikom broju efektornih ćelija, uključujući i glatkih mišića, sekretornih ćelija (šef, parijetalni, sluz, enterocytes, gušterače egzokrine ćelija) i gastrointestinalnog endokrine ćelije.
Interneurona su u velikoj mjeri odgovorni za integraciju informacija iz senzornih neurona i pružanje ga (“programiranje”) crijevne motornih neurona.
Crijevne neuroni luče zastrašujuće niz neurotransmitera. Jedan od glavnih neurotransmitera u produkciji crijevne neurona je acetilkolin. U principu, neuroni koji luče acetilkolina su eksitatorne, stimulisanje kontrakcije glatkih mišića, povećanje crijevnih sekretu, oslobađanje crijevne hormona i širenje krvnih žila. Norepinefrin se koristi intenzivno za neurotransmitera u probavnom traktu, ali to proizlazi iz spoljašnji simpatičkog neurona; učinak noradrenalina je gotovo uvijek inhibitorni i suprotno da acetilkolina.
Na crijevne nervni sistem može i funkcioniše samostalno, ali normalnu funkciju probavnog zahtijeva komunikacijske veze između ovog unutrašnjeg sistema i centralnog nervnog sistema. Ovi linkovi imaju oblik parasimpatičkog i simpatičkog vlakana koje povezuju ili centralnog i crijevne nervnog sistema ili povezivanje centralnog nervnog sistema direktno sa probavnog trakta. Kroz ove cross veze, crijevima može pružiti senzornih informacija do CNS-a CNS mogu utjecati na gastrointestinalni funkciju. Priključak na centralni nervni sistem također znači da signali izvan digestivnog sistema može se prenose na probavni sistem: na primjer, prizor privlačan hrane stimulira lučenje u želucu.
U principu, simpatičke stimulacije uzrokuje inhibiciju gastrointestinalnog sekrecije i motoričke aktivnosti, i skupljanje gastrointestinalnog sfinktera i krvnih sudova. Nasuprot tome, parasimpatički podražaja obično stimuliše ove aktivnosti probavnog. Neki od istaknutih saopštenjima omogućena nervozan interkonekcija unutar probavnog trakta su imenovani kao reflekse i služe za ilustraciju robustan sistem kontrole. Primjeri uključuju gastrocolic refleks, gdje distention želuca stimuliše evakuaciju debelog crijeva, a enterogastric refleks, u kojem distention i iritaciju tankog crijeva dovodi do supresije lučenja i motoričke aktivnosti u želucu.
Kongenitalne i stečene poremećaj u strukturi ili funkciji crijevne nervnog sistema i prepoznati kao uzroka bolesti probavnog trakta. Primjeri uključuju male crijevnih poremećaja motiliteta, želuca utičnicu prepreke i megakolon.
Otkriće da su ptice evoluirale od malih dinosaura mesoždera iz kasne jure je omogućeno nedavno otkrili fosile iz Kine, Južne Amerike i drugih zemalja, kao i gledajući stare muzej uzoraka iz nove perspektive i sa novim metodama. Potraga za preci žive ptica počela je primjerak Archaeopteryxa, prvi poznati ptica, otkriven u ranim 1860-ih. Kao ptice, imao je perje duž svoje ruke i rep, ali za razliku od žive ptice, takođe je zube i dug koščati rep. Osim toga, mnogi od kostiju u Archaeopteryxaruke ‘s, rame pojaseva, zdjelice i stopala bila jasna, nije spojena i smanjeni su u žive ptice. Na osnovu ovih karakteristika osnovu Archeopteryx je prepoznat kao srednji između ptica i reptila; ali koji reptili?
Berlinski primerak Archaeopteryxa lithographica
U 1970-ih, paleontolozi primijetio da Archaeopteryx dijele jedinstvene karakteristike s malim mesoždera dinosaurusa zove teropoda. Sve dinosaurusa grupe na ovom evogram, osim Ornithischia dinosaurusi su teropoda. Na njihove zajedničke osobine zasnovane, naučnici obrazložio da je možda teropodi su preci ptica. Kada paleontolozi izgrađen evolucijski drveća da prouči pitanje, oni su još više uvjereni. Ptice su jednostavno grančica na grani dinosaurusi “stabla života.
Kao što su ptice evoluirale od ovih teropoda, mnoge njihove karakteristike su modifikovani. Međutim, važno je imati na umu da su životinje ne “pokušavamo” biti ptica u bilo kojem smislu. U stvari, bliže gledamo, više očito je da je paket osobina koje karakteriziraju ptice evoluirale kroz kompleksni niz koraka i služio različitim funkcijama usput.
Uzmite perje, na primjer. Mali teropoda koji se odnose na Compsognathus (npr Sinosauropteryx ) vjerojatno evoluirala prvi perje. Ove kratke, kose poput pera porastao na glavama, vrata i tijela i pruža izolaciju. Perje Izgleda da su imali različite obrasce boja, kao i, iako bez obzira da li su to bili za prikaz, maskirne, prepoznavanje vrste, ili neku drugu funkciju je teško reći.
Ovaj fosil Sinosauropteryx čuva dokaza perja kose nalik.
U teropodi još više usko vezana za ptice, poput oviraptorosaurs, nalazimo nekoliko novih tipova perja. Jedan je razgranata i plamenjača, kao na slici ispod. Drugi su evoluirali centralni stabljika, s nestrukturiranim grane koje izlaze je i svoju bazu. Ipak drugi (poput dromaeosaurids i Archaeopteryx ) imaju vane nalik strukturu u kojoj su bodlje su dobro organizovani i zaključao zajedno barbules. Ovo je identična struktura perja žive ptice.
U redu, asimetrične perje let su prisutni u fosil dromaeosaurid koji je možda imao mogućnost da klizi.
Druga linija dokaza dolazi iz promjena u cifre dinosaurusa vodi do ptice. Prvi teropoda imao ruke sa malim peti i četvrti cifre i dug druga cifra. Kao što evogram pokazuje, u teropoda lozi koja će na kraju dovesti do ptice, peta cifra (npr kao što se vidi u Coelophysoids), a zatim četvrti (npr kao što se vidi u alosauridima) je potpuno izgubljen. Kosti zgloba osnovi prvog i drugog cifre konsolidovali i odveo na polukružni oblik koji je omogućio ruke na stranu okretati protiv podlaktice. Krilo spojevi ovog eventualno dozvoljeno ptica ‘za kretanje na način koji stvara potisak za let.
Ovo oviraptorid dinosaurusa, Citipati osmolskae , možda štiti gnijezdo jaja.
Funkcije perje kao što su evoluirali su dugo raspravlja. Kao što smo vidjeli, prvo najjednostavniji, kosa nalik na perje očito služio kao insulatory funkciju. Ali u kasnijim teropoda, kao što su neki oviraptorosaurs, perje na ruke i šake su dugo, iako su i sami Prednji udovi su kratki. Šta su ti životinje s dugim perjem na kratko oružje? Jedna sugestija dolazi iz neke izuzetne fosili oviraptorosaurs čuva u krede sedimentima pustinje Gobi. Skelet životinja nagnut se na gnijezdo jaja, kao zamišljen piletine. Ruke su rašireni jaja kao da ih kriju. Dakle, možda ove perje je bilo u funkciji zagrijavanja jaja i štiti ih od zla.
Ptice nakon Archeopteryx nastavio razvija u nekim istom pravcu kao svoju theropodnih predaka. Mnogi od njihove kosti su smanjeni, a spojene, koja je možda pomogla povećati efikasnost leta. Slično tome, kosti zidovi postala još tanji, a perje je postao duži i njihove lopatice asimetrične, vjerovatno i poboljšanje let. Koštanog rep je sveden na panj, i sprej perja na repu na kraju je uzeo na funkciji poboljšanja stabilnosti i upravljivost. Jadac, koja je bila prisutna u dinosaurusi nisu ptica, postao je jači i složenije, i kosti ramenog obruča evoluirao kako bi se povezali sa grudne kosti, sidrenje aparat let forelimb. Sama grudne kosti postaju veći, i evoluirao centralni kobilica duž središnje linije dojke koja je služila za sidro mišiće let. Ruke evoluirala da bude duži od nogu, kao glavni oblik kretanja koji prebacio iz trči prema leta, a zubi su više puta izgubio u raznim loze ranoranioci. Predak svih živih ptica živio negdje u kasnom krede, au 65 miliona godina od izumiranje ostatka dinosaurusa, ovaj predaka preinačen u glavne grupe ptica danas živ.
Somatostatin je prvi put otkriven u hipotalamusa ekstrakti i identificirana kao hormon koji inhibira lučenje hormona rasta. Nakon toga, somatostatin je utvrđeno da se luči širok spektar tkiva, uključujući i gušterača, trakta i regiona centralnog nervnog sistema van hipotalamusa.
Struktura i sinteza
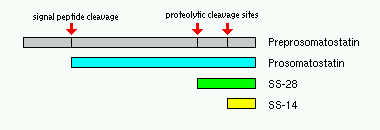
Dva oblika somatostatin sintetiziraju. Oni se nazivaju SS-14 i SS-28, što je odraz njihove amino dužina kiselina lanca. Oba oblika somatostatin generiraju proteolitičke dekolte od prosomatostatin, koji je i sam potiče iz preprosomatostatin. Dva cistein residules u SS-14 omogućavaju peptida da se formira interne disulfid obveznica.
Relativne količine SS-14 u odnosu na SS-28 luči zavisi tkiva. Na primjer, SS-14 je dominantan oblik proizvedenog u nervnog sistema i očito jedini oblik luči iz pankreasa, dok je crijevo luči uglavnom SS-28.
Osim razlike tkiva specifična u lučenju SS-14 i SS-28, dva oblika ovog hormona može imati različite biološki potencija. SS-28 je otprilike deset puta moćniji u inhibicije hormona rasta sekrecije, ali manje potentni da SS-14 u inhibicije glukagon oslobađanje.
Receptori i Mehanizam djelovanja
Pet stomatostatin receptori su identificirani i karakteriše, od kojih su svi članovi G protein spregnutih receptora natporodice. Svaki od receptora aktivira različite mehanizme signalizaciju u ćelijama, iako su svi inhibiraju adenilil ciklazu. Četiri od pet receptora ne prave razliku SS-14 iz SS-28.
Fiziološki efekti
Somatostatin akata i endokrinih i parakrinu puteve da utiču na svoje ciljne ćelije. Pojavljuje se većina kruži somatostatin da dođe iz pankreasa i gastrointestinalnog trakta. Ako se morao sumirati efekte somatostatin u jednu rečenicu, to bi bilo: “somatostatin inhibira izlučivanje mnogih drugih hormona “.
1.Efekti na hipofizi
Somatostatin je imenovan za učinak inhibicije sekrecije hormona rasta iz hipofize. Eksperimentalno, sve poznate stimulanse za hormon rasta lučenje su potisnuti somatostatin administracije. Pored toga, životinja tretiranih antiseruma na somatostatin pokazuju povišene koncentracije u krvi hormona rasta, kao i životinje koje su genetski modifikovane poremetiti njihove somatostatin gena.
Na kraju krajeva, hormon rasta lučenje je pod kontrolom interakcije somatostatin i hormona rasta oslobađanje hormona, od kojih su oba luče hipotalamusa neurona.
2.Efekti na pankreas
Ćelija u pankreasa otočića luče inzulin, glukagon i somatostatin. Izgleda somatostatin prvenstveno djelovati na parakrini način da inhibira sekreciju inzulin i glukagon. Ona također ima učinak u suzbijanju pankreasa egzokrine sekreta, inhibirajući holecistokinina -stimulisano enzima lučenje i sekretinska – stimulisano bikarbonata sekrecije.
Pored direktnih efekata inhibicije lučenja drugih GI hormona, somatostatin ima niz drugih inhibira GI trakta, što može odražavati njegove efekte na druge hormone, plus neke dodatne direktne efekte. Somatostatin potiskuje lučenje želučane kiseline i pepsina, smanjuje stopu pražnjenje želuca i smanjuje glatke mišićne kontrakcije i protok krvi u crijevima. Zajedno, ove aktivnosti izgleda da imaju ukupni efekt smanjenja stope apsorpcija nutrijenata.
4.Efekti na nervni sistem
Somatostatin se često naziva kao da ima neuromodulatornih aktivnost u okviru centralnog nervnog Sytem, a čini se da imaju različite složene efekte na neuralne transmisije. Ubrizgavanje somatostatin u mozak glodavaca dovodi do stvari kao što su povećana uzbuđenje i smanjenje sna, i umanjenje vrijednosti nekih motornih odgovora.
Farmakološke upotreba somatostatina
Somatostatin i njegove sintetički analozi koriste se klinički za liječenje raznih tumora. Također se koristi u za liječenje gigantizam i akromegalija, zbog svoje sposobnosti da inhibiraju rast lučenje hormona.
Oko 60% mase tijela je voda, i bez obzira na velike varijacije u iznosu od vode uzima u svaki dan, sadržaj vode u telu ostaje izuzetno stabilna. Takva preciznu kontrolu koncentracije tijela vode i rastvorene je funkcija nekoliko hormona djeluje na oba bubrega i vaskularnog sistema, ali nema sumnje da antidiuretskog hormona je ključni igrač u ovom procesu.
Antidiuretskog hormona, poznat i često kao arginin vazopresin, je kiselina peptida devet amino luči iz zadnjeg hipofize. Unutar hipotalamusa neurona, hormona je pakiran u sekretorni vezikule s nosača protein neurophysin, a oba su pušteni na lučenje hormona.
Fiziološki efekti Antidiuretski hormona
Efekti na bubrege
Najvažniji efekat antidiuretskog hormona je očuvanje vodnog tijela smanjujući gubitak vode u urinu. A diuretik je agent koji povećava brzinu formiranja urina. Ubrizgavanje malih količina antidiuretskog hormona u rezultate osobe ili životinje u antidiuresis ili smanjena formiranja urina, a hormon je imenovan za ovaj efekat.
Antidiuretskog hormona se vezuje za receptore na ćelijama u sabirnih kanala bubrega i promovira reapsorpcije vode natrag u cirkulaciju. U absense od antidiuretskog hormona, sabirnih kanala praktično nepropustljivim na vodu, i to ističe kao urin.
Antidiuretskog hormona stimulira vode reabsorbtion stimulirajući umetanje “vode kanali” ili akvaporini u membrane tubula bubrega. Ovi kanali transport rastvorene bez vode kroz cjevaste ćelije i natrag u krv, što je dovelo do smanjenja u plazmi osmolarnost i povećanje osmolarnost urina.
Efekti na vaskularni sistem
Kod mnogih vrsta, visoke koncentracije antidiuretskog hormona izazvati široko rasprostranjene suženje arteriola, što dovodi do povećanja arterijskog pritiska. Bilo je to za ovaj efekat da je ime vazopresin skovan. U zdravih ljudi, antidiuretskog hormona ima minimalne efekte presorsku.
Kontrola Antidiuretski lučenje hormona
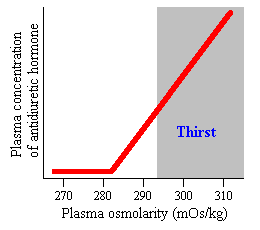
Najvažniji varijabla regulišu antidiuretskog hormona je plazma osmolarnost, ili koncentraciju rastvorenih materija u krvi. Osmolarnost se oseti u hipotalamusu strane neurona poznat kao osmoreceptors, a oni neuroni, zauzvrat, stimulišu lučenje iz neurona koji proizvode antidiuretskog hormona.
Kada plazma osmolarnost ispod određenog praga, osmoreceptors se ne aktivira i secretio od antidiuretskog hormona je potisnut. Kada osmolarnost povećava iznad praga, stalno popustu osmoreceptors ovo prepoznaju kao svoj znak da stimulišu neuroni koji luče antidiuretskog hormona. Kao što se vidi na slici ispod, antidiuretskog koncentracije hormona naglo rastu i linearno sa povećanjem plazma osmolarnost.
Osmotski kontrolu nad antidiuretskog hormona ima smisla. Zamislite hodanje preko pustinje: sunce tuče dolje i počnete izgubiti znatnu količinu tjelesne vode kroz znojenje. Gubitak rezultati vode u koncentraciji krvi rastvorenih materija – plazma osmolarnost povećava. Treba da povećaju proizvodnju urina u takvoj situaciji? Jasno da ne. Umjesto toga, antidiuretskog hormona se luči, što omogućava gotovo svu vodu koja bi se izgubili u urinu da se resorbuje i konzervirani.
Tu je zanimljiva paralela između antidiuretskog hormona i žeđ. Oba fenomena Čini se da je stimuliše hipotalamusa osmoreceptors, iako vjerojatno ne iste. Osmotski prag za antidiuretskog hormona je znatno niža nego za žeđ, kao da hipotalamus se kaže “Hajde da ga ne smeta pozivajući žeđ, osim ako je situacija toliko loša da antidiuretskog hormona ne mogu nositi sami.”
Lučenje antidiuretskog hormona takođe stimuliše smanjenje krvnog pritiska i zapremine, uslovi očitava protežu receptore u srcu i velikim arterijama. Promjene krvnog pritiska i volumena nisu ni približno kao osjetljiva stimulator i povećanje osmolarnost, ali su ipak moćni u teškim uslovima. Na primjer, gubitak od 15 ili 20% volumena krvi rezultati krvarenje u masivnom sekrecije antidiuretskog hormona.
Još jedan moćan stimulans antidiuretskog hormona je mučnina i povraćanje, a oba su pod kontrolom područja u mozgu s linkovima na hipotalamus.
Stablo stanja
Najčešće bolesti čovjeka i životinja koje se odnose na antidiuretskog hormona je dijabetes insipidus. Ovo stanje može nastati iz bilo koje od dvije situacije:
Hipotalamusa (“Central”) dijabetes insipidus je rezultat nedostatak u sekrecije antidiuretskog hormona iz zadnjeg hipofize. Uzroci ove bolesti uključuju povrede glave, i infekcija ili tumora uključuje hipotalamus.
Nefrogene dijabetes insipidus nastaje kada bubreg nije u stanju da odgovori na antidiuretskog hormona. Najčešće, to proizlazi iz neke vrste bolesti bubrega, ali mutacije receptora gena ADH ili gena koji kodira AQUAporin-2 su i pokazali u ugroženim ljudima.
Glavni znak oba tipa dijabetesa insipidus prevelika proizvodnja urina. Neki ljudski pacijenti proizvoditi čak 16 litara urina dnevno! Ako je dostupna za potrošnju adekvatnu vode, bolest je rijetko opasne po život, ali uskraćivanje vode može biti vrlo opasno. Hipotalamusa dijabetes insipidus mogu biti tretirani sa egzogenim antidiuretskog hormona.
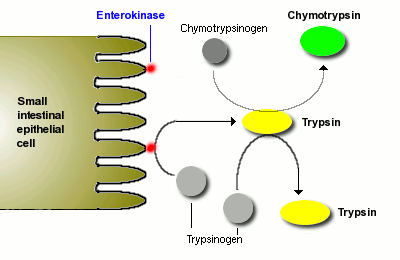
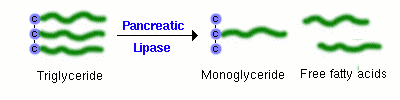
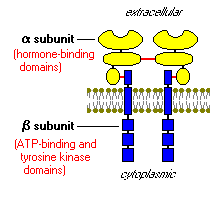
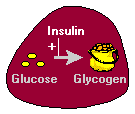
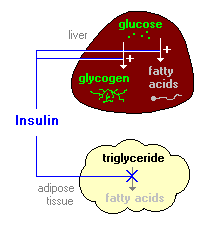


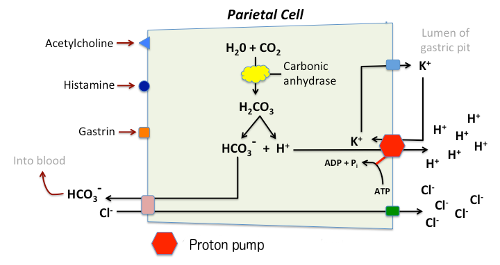

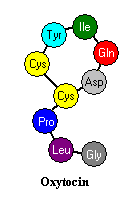
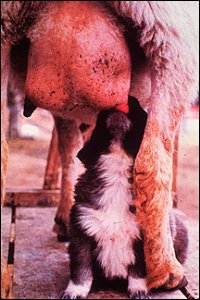

 Merozoites brzo (oko 20 sekundi) i posebno ući eritrocita.Ova specifičnost se ogleda i za eritrocita kao preferirani tip ćelije domaćina i za određenu vrstu domaćina, implicirajući receptor-ligand interakcije.Eritrocita invazija je složen proces koji se samo djelomično shvatiti na molekularnom i ćelijskom nivou. Međutim, značajan napredak je postignut u identifikaciji mnogih parazita i domaćina proteina koji su važni za proces invazije.
Merozoites brzo (oko 20 sekundi) i posebno ući eritrocita.Ova specifičnost se ogleda i za eritrocita kao preferirani tip ćelije domaćina i za određenu vrstu domaćina, implicirajući receptor-ligand interakcije.Eritrocita invazija je složen proces koji se samo djelomično shvatiti na molekularnom i ćelijskom nivou. Međutim, značajan napredak je postignut u identifikaciji mnogih parazita i domaćina proteina koji su važni za proces invazije. Sadržaj apeksnog organela su protjerani kao parazit napadne, sugerišući da su ove organele igraju neku ulogu u invaziji. Eksperimenti u Toxoplasma gondii ukazuju na to da je micronemes su protjerani prvi i doći sa inicijalni kontakt između parazita i domaćina (
Sadržaj apeksnog organela su protjerani kao parazit napadne, sugerišući da su ove organele igraju neku ulogu u invaziji. Eksperimenti u Toxoplasma gondii ukazuju na to da je micronemes su protjerani prvi i doći sa inicijalni kontakt između parazita i domaćina (
 Još jedna porodica adhesins uključenih u vezivanju merozoites na erthrocytes su retikulocita vezivanja poput kolegama (RH).Članovi ove porodice udjela homologije s protein prvobitno identifikovan u P. vivax koji se vezuje specifično retikulocita i mogu igrati ulogu u retikulocita specifičnosti P. vivax. Ostali microneme proteina u ‘TRAP’ porodice su također uključeni u lokomotorni i/ili ćelija invaziju na pozornici sporozoite i drugih apicomplexa (
Još jedna porodica adhesins uključenih u vezivanju merozoites na erthrocytes su retikulocita vezivanja poput kolegama (RH).Članovi ove porodice udjela homologije s protein prvobitno identifikovan u P. vivax koji se vezuje specifično retikulocita i mogu igrati ulogu u retikulocita specifičnosti P. vivax. Ostali microneme proteina u ‘TRAP’ porodice su također uključeni u lokomotorni i/ili ćelija invaziju na pozornici sporozoite i drugih apicomplexa ( Ova interakcija je dodatno ojačana oslobađanje dva dodatna protein kompleksa iz regiona vrat rhoptries (
Ova interakcija je dodatno ojačana oslobađanje dva dodatna protein kompleksa iz regiona vrat rhoptries (



 Pf EMP1 je član var gena porodice (
Pf EMP1 je član var gena porodice ( Kodiranje cytoadherence ligand od strane visoko polimorfne gen porodice predstavlja paradoks u tom
Kodiranje cytoadherence ligand od strane visoko polimorfne gen porodice predstavlja paradoks u tom